全國服務咨詢熱線:
將體外轉錄的 RNA 用作治療藥物需要大量具有低免疫原性的功能性 RNA。用于合成這些體外轉錄的 mRNA 的技術——主要使用 T7 噬菌體 RNA 聚合酶 (T7 RNAP),這個技術目前已經很成熟。T7 RNAP 從包含噬菌體酶特異性啟動子的 DNA 模板中以高保真度轉錄 RNA。盡管使用 T7 RNAP 從 DNA 模板合成 RNA 的過程很穩健,但之前的研究已經確定了體外合成過程中某些副產物的存在,這些副產物會觸發細胞免疫反應,包括雙鏈 RNA (dsRNA),這已被證明成為免疫通路的主要觸發因素。因此,在為尋求最小化細胞免疫反應的體內應用合成 mRNA 時,從 mRNA 制劑中消除這些 dsRNA 污染物或減少它們的形成是至關重要的。

dsRNA 副產物的產生被認為主要通過兩種不同的機制發生。首先,由 T7 RNAP 合成的 RNA 轉錄本(runoff轉錄本,圖1)在后續幾輪轉錄中作為 T7 RNAP 的 RNA 依賴性 RNA 聚合酶活性的模板[1]。如果runoff轉錄本的 3'-末端具有足夠的互補性(在順式中),它將向后折疊并導致runoff轉錄本的延伸。生成的 RNA 將在 3' 端延伸,并且可以在凝膠上的變性條件下與主要轉錄物區分開來(圖2A)。
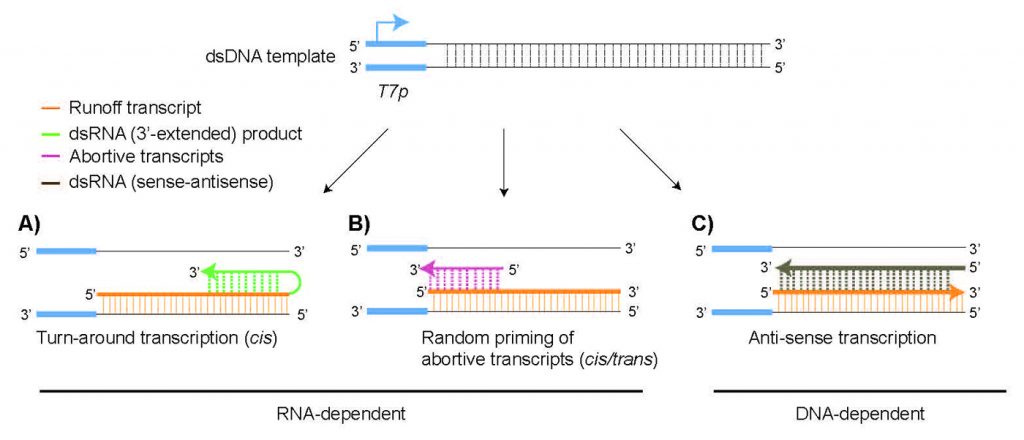
圖1 體外轉錄過程中 dsRNA 副產物形成的可能機制示意圖
短的 RNA 片段,例如流產的轉錄物,可以退火到runoff轉錄物中的互補序列(反式),并且還會導致 dsRNA 副產物的形成(圖1B)。dsRNA 產物的身份和性質將根據延伸發生在順式還是反式而有所不同,并且預計 RNA 積累與虛假產物的形成之間存在很強的相關性,因為合成的 RNA可能會重新結合聚合酶以啟動延伸。
第二種dsRNA 副產物機制是RNAP 可能會切換到非模板鏈,從而產生與runoff產物互補但以啟動子獨立方式合成的 RNA 分子(圖1C)[2]。由于反義分子的大小與runoff產物的大小相似,因此無法通過變性凝膠電泳進行區分。相反,分析由于存在反義 RNA 分子而形成的 dsRNA 副產物將需要天然條件(圖2B)。
1)凝膠電泳法
以runoff轉錄本為模板、順式或反式形成的dsRNA,其長度短于runoff轉錄本,在變性凝膠下可與目標轉錄本區分(圖2A)。以DNA模板或非模板鏈為模板轉錄生成的runoff轉錄本或反義鏈,二者結合形成的dsRNA長度與runoff轉錄本一致,無法通過變性凝膠有效分離,可使用活性膠實現dsRNA與ssRNA的分離(圖2B)。兩種凝膠電泳法適用于不同副產物的定性檢測,分辨率相對較低,不能精確地定量分析dsRNA殘留量。
2)免疫印跡法
免疫印跡法利用dsRNA特異性抗體來檢測IVT反應中dsRNA含量。J2抗體是一種抗dsRNA抗體,可特異性識別并結合dsRNA,根據特定信號鑒別樣品中是否存在dsRNA(定性檢測),如圖2C所示。同時制備合適的標準對照品,以定量分析待測樣品的dsRNA殘留,提高檢測的靈敏度(圖2E)。該方法依賴于抗體對dsRNA的特異性識別作用,而mRNA二級結構或修飾核苷酸可能改變抗體對dsRNA結構的識別,需進行專門的研究。注意:抗體檢測的靈敏度與dsRNA區域的長度有關,需調整檢測的靈敏度以確保dsRNA可被檢測。
3)酶法免疫印跡法
是當前dsRNA檢測的常用方法,可采用尼龍膜或商業化酶聯免疫法(ELISA)試劑盒。使用尼龍膜上樣需優化上樣體積,以使各樣品擴散程度一致,減少對結果精確度的影響。商業化試劑盒檢測過程相對簡單。以某ELISA試劑盒為例,基于雙抗體夾心酶聯免疫法,使用捕獲抗體包被微孔板,形成固相抗體,向固相抗體微孔板中加入dsRNA校準品和待測樣品,然后加入檢測抗體,最后加入辣根過氧化物酶(HRP)標記的酶標二抗,形成“包被抗體-dsRNA-酶標檢測抗體"復合物,經過洗滌后加入顯色液顯色,終止反應后使用酶標儀檢測吸光值,吸光值與樣品中dsRNA的量呈相關性。
4)MDA5特異結合法
胞質免疫受體MDA5是一種RIG-I樣受體(RIG-I like receptors,RLRs),可特異性識別dsRNA從而激活機體的抗病毒免疫反應,因此可根據MDA5與dsRNA的結合作用設計體外檢測策略。首先,使用冷凍電子顯微鏡(cryo-EM)可觀察到MDA5-dsRNA復合體存在短纖絲(filaments),對短纖絲結構的分析顯示,每個MDA5分子橫跨14-15個RNA堿基對。但這一分析方法因低溫 EM 分析的設備要求而受到限制。其次,MDA5是一種ATP依賴性解旋酶,MDA5與dsRNA的結合可激活MDA5的腺嘌呤核苷三磷酸(ATP)水解酶活性,可可以通過標準生化測定法檢測dsRNA。因此,在存在 MDA5 的情況下進行 ATP 水解的體外測定可能是了解目標 mRNA 產生的免疫反應程度的良好替代方法。
5)測序或質譜分析
為深入研究dsRNA的核苷酸序列,可進行RNA-seq2或完整分子的質譜分析(圖2D)。兩種方案的不足之處在于,RNA-seq分析過程中結構化RNA的連接偏倚可能影響實驗結果,而質譜分析提供所選RNA的異質性分布,但無法提供RNA 3’端的序列信息。因此,兩種分析方法并不通用,且在篩選多個序列時不能以高通量的方式進行。
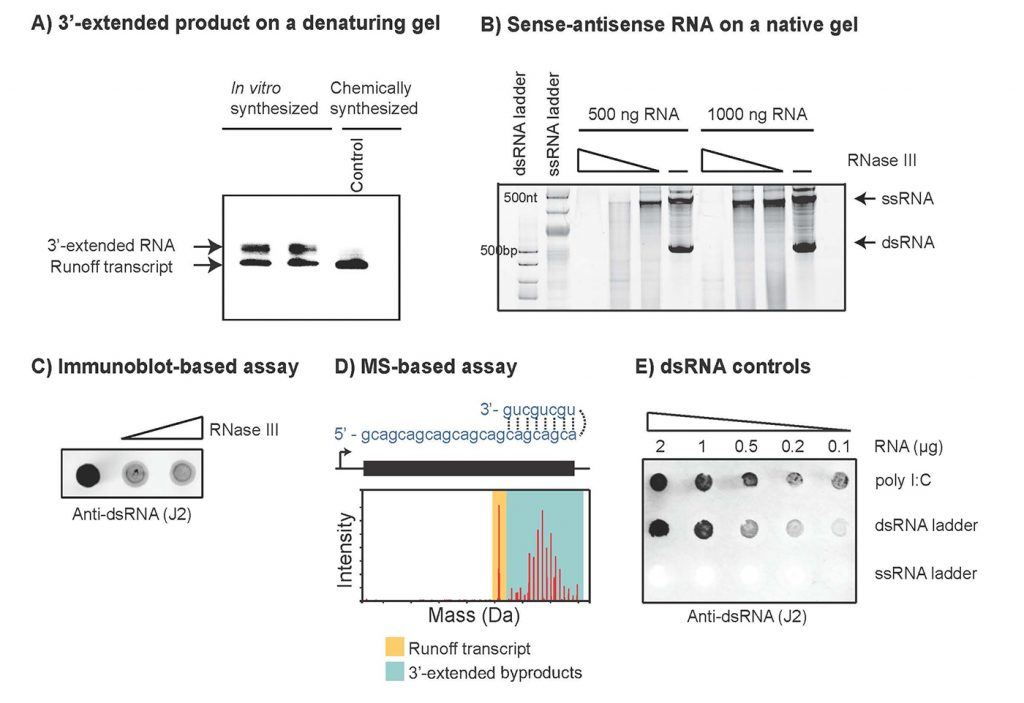
由于免疫測定無法區分 dsRNA 產物是由于 3' 端延伸還是反義分子合成而形成,因此建議實施不止一種方法來表征副產物的性質,因為了解分子的性質可以指導采取何種方法來防止在 IVT 反應中形成 dsRNA 副產物(見下一節)。
1)高效液相色譜 (HPLC) 純化
高效液相色譜 (HPLC) 的純化已被證明可以將 dsRNA 副產物與主要 IVT RNA 產物分離(圖 3A)[3]。HPLC 純化過程中 dsRNA 副產物的保留時間較長表明該方法可用于將 3'-擴展的 dsRNA 副產物與主要 IVT RNA 分離。這種方法雖然有效,但會導致 mRNA 合成工作流程中的額外步驟,涉及專門的儀器,與擴大反應不兼容,并阻礙了該方法的成本效益。
此外,尚不清楚這種基于 HPLC 的方法是否可以有效分離反義 dsRNA 產物,它們的分離可能取決于所使用的實驗條件。還報道了基于 dsRNA 與纖維素的選擇性結合,用于去除 dsRNA 污染物的基于纖維素的色譜方法[4]。盡管這種純化策略具有成本效益,而且這種方法的一次性性質可以防止先前純化的殘留,但它可能并不適合所有 mRNA 序列,因為某些 mRNA 可能更傾向于形成二級結構這可能會降低恢復率。
2)使用耐熱 RNA 聚合酶進行IVT 反應
合成后純化的另一種方法是通過改變 IVT 反應條件來防止在中形成 dsRNA 副產物。降低 IVT 反應中的鎂含量以減少一些特定模板[2]的 dsRNA 副產物(通過反義 RNA 的合成形成)的形成;然而,降低反應中的鎂濃度也會影響 RNA 的總產量,這對于需要大量 mRNA 的應用來說是不可取的。
最近表明,添加與runoff轉錄本 3' 端互補的 DNA 寡核苷酸也可以防止 3' 延伸的 dsRNA 副產物的形成[5],但在合成治療性 mRNA 后去除這種寡核苷酸可能具有挑戰性,并且需要額外的酶促步驟,這在試圖簡化工作流程和限制生產成本時是不可取的。
使用耐熱 RNA 聚合酶,例如來自 New England Biolabs 的 HiT7® 已被證明可以減少 IVT 反應中 3'-延伸的 dsRNA 副產物(圖 3B)的形成,而不影響總產量或RNA,并且它不需要額外的酶處理,這可以為當前的 IVT 反應工作流程提供替代方案[6, 7]。
適合去除/預防 dsRNA 污染物的方法將取決于最終應用和所需的 RNA 產量規模。對于以放大為先決條件的應用,合成后純化步驟可能會阻礙最終結果。更好的方法是防止在合成過程中形成 dsRNA 副產物。
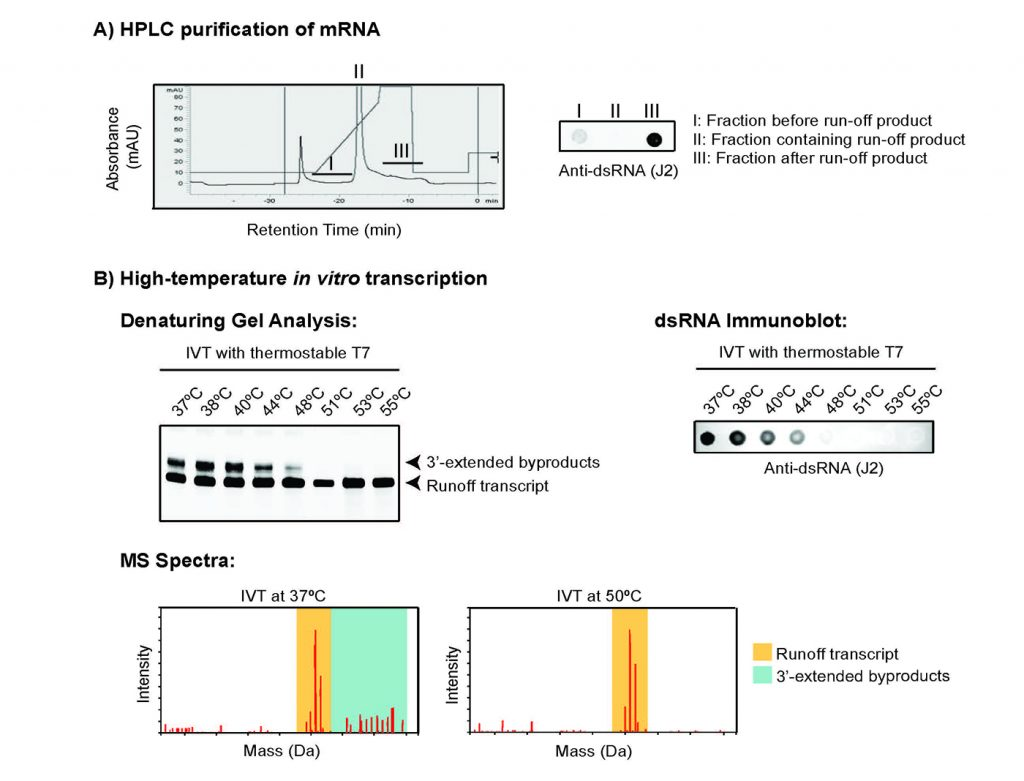
圖3 減少IVT 反應中 dsRNA 副產物。
使用合成 mRNA 作為藥物需要 mRNA 不含任何污染 RNA,并且需要大量合成。因此,在選擇合適的方法去除 dsRNA 副產物以合成您喜歡的 mRNA 分子時,應盡早考慮 IVT 反應的最終應用和規模。dsRNA 副產物也是必須考慮的一個因素。例如,模板編碼的 poly-A 加尾可以避免部分 dsRNA 副產物的形成,但不是全部,高溫轉錄可以減少 3'-末端延伸的 dsRNA 副產物,但不能減少反義 RNA 的合成[7]。使用高產率反應條件來合成更大量的 RNA 對于需要放大的應用來說很有吸引力。然而,由于在存在過量 RNA 的情況下可以增強某些 dsRNA 副產物的形成,因此應考慮酶:模板:NTP 條件,并針對每個序列進行優化,盡可能地減少這些 dsRNA 副產物的形成。DNA 模板或 RNA 分子中更容易形成 dsRNA 副產物的序列的性質尚不清楚。
更好地了解序列特異性有助于 mRNA 3'-末端的合理設計,以防止在反應中形成這些污染物。即使是模板序列和/或反應條件的微小變化也可能影響 dsRNA 副產物形成的程度,在設計/修改模板序列時應予以考慮。最后,在使用經過修改的 NTP 制作 mRNA 時,很難檢測 dsRNA 副產物,在這種情況下,至少應采用一種以上的方法來表征和定量 dsRNA 副產物。
總之,用于治療目的的合成 mRNA 的高效且具有成本效益的生產將需要對最終 mRNA 產品的性質有透徹的了解,并且將取決于能夠最大限度地減少污染物產生的合成過程,否則將需要昂貴的純化方法。
參考文獻
【1】 Gholamalipour Y, Karunanayake Mudiyanselage A, Martin CT. 3’ end additions by T7 RNA polymerase are RNA self-templated, distributive and diverse in character-RNA-Seq analyses. Nucleic Acids Res. 2018 Oct 12;46(18):9253-9263.
【2】 Mu X, Greenwald E, Ahmad S, Hur S. An origin of the immunogenicity of in vitro transcribed RNA. Nucleic Acids Res. 2018 Jun 1;46(10):5239-5249.
【3】 Karikó K, Muramatsu H, Ludwig J, Weissman D. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. 2011 Nov;39(21):e142.
【4】 Baiersd?rfer M, Boros G, Muramatsu H, Mahiny A, Vlatkovic I, Sahin U, Karikó K. A Facile Method for the Removal of dsRNA Contaminant from In Vitro-Transcribed mRNA. Mol Ther Nucleic Acids. 2019 Apr 15;15:26-35.
【5】 Gholamalipour Y, Johnson WC, Martin CT. Efficient inhibition of RNA self-primed extension by addition of competing 3’-capture DNA-improved RNA synthesis by T7 RNA polymerase. Nucleic Acids Res. 2019 Aug 8. pii: gkz700. doi: 10.1093/nar/gkz700.
【6】 Roy B, Robb GB. Use of thermostable RNA polymerases to produce RNAs having reduced immunogenicity. US Patent 10,034,951.
【7】 Wu MZ, Asahara H, Tzertzinis G, Roy B. Synthesis of low immunogenicity RNA with high-temperature in vitro transcription.
酶產品
貨號 | 產品名稱 | 規格 |
abs60153 | T7 RNA 聚合酶 | 50KU/200KU/1000KU |
工具RNA
貨號 | 產品名稱 | 規格 |
abs60176 | EGFP mRNA | 100ug/100ug×5/100ug×10 |
abs60178 | Fluc mRNA | 100ug/100ug×5/100ug×10 |
abs60180 | mcherry mRNA | 100ug/100ug×5/100ug×10 |
abs60191 | Cre mRNA | 100ug/100ug×5/100ug×10 |
abs60312 | OVA mRNA | 100ug/100ug×5/100ug×10 |
abs60174 | Cas9 mRNA | 100ug/100ug×5/100ug×10 |
abs60192 | EBFP mRNA | 100ug/100ug×5/100ug×10 |
abs60193 | ECFP mRNA | 100ug/100ug×5/100ug×10 |
abs60332 | Endless eGFP mRNA | 50ug |
abs60333 | Endless Fluc mRNA | 50ug |
abs60334 | Endless mcherry mRNA | 50ug |
abs60335 | Endless Cas 9 mRNA | 50ug |
abs60336 | Endless EPO mRNA | 50ug |
核酸電泳產品
貨號 | 產品名稱 | 規格 |
abs9380 | BIS-Tris Gels 預制膠4-12%, 10 wells | 10片/盒 |
abs9381 | BIS-Tris Gels 預制膠4-12%, 12 wells | 10片/盒 |
abs9382 | BIS-Tris Gels 預制膠4-12%, 15 wells | 10片/盒 |
abs9383 | BIS-Tris Gels 預制膠4-20%, 10 wells | 10片/盒 |
abs9384 | BIS-Tris Gels 預制膠4-20%, 12 wells | 10片/盒 |
abs9385 | BIS-Tris Gels 預制膠4-20%, 15 wells | 10片/盒 |
abs9386 | BIS-Tris Gels 預制膠8%, 10 wells | 10片/盒 |
abs9387 | BIS-Tris Gels 預制膠8%, 12 wells | 10片/盒 |
abs9388 | BIS-Tris Gels 預制膠8%, 15 wells | 10片/盒 |
abs9389 | BIS-Tris Gels 預制膠10%, 10 wells | 10片/盒 |
abs9390 | BIS-Tris Gels 預制膠10%, 12 wells | 10片/盒 |
abs9391 | BIS-Tris Gels 預制膠10%, 15 wells | 10片/盒 |
abs9392 | BIS-Tris Gels 預制膠12%, 10 wells | 10片/盒 |
abs9393 | BIS-Tris Gels 預制膠12%, 12 wells | 10片/盒 |
abs9394 | BIS-Tris Gels 預制膠12%, 15 wells | 10片/盒 |
ELISA檢測相關試劑盒
貨號 | 產品名稱 | 規格 |
abs590002 | 雙鏈RNA(dsRNA)ELISA檢測試劑盒(K1) | 96T |
Absin特色產品線:
WB相關:ECL發光液、預染marker、預制膠;IHC相關:二抗試劑盒、組化筆;IP/CoIP試劑盒;激動劑/抑制劑;血清、BSA、蛋白酶K、CTB、TTX、CEE;凋亡試劑盒;呼吸爆發試劑盒;ELISA試劑盒;重組蛋白;抗體: 二抗、標簽抗體、對照抗體;定制服務(抗體/多肽/蛋白/標記/檢測)...
電話
微信掃一掃
